







au sommaire
Un projet ambitieux entièrement privé : le projet génographique. Établir l'arbrearbre généalogique de l'humanité et en déduire l'expansion des hominidéshominidés sur la planète. C'est l'étude des ADN (acide désoxyribonucléiqueacide désoxyribonucléique) de nos contemporains qui permettra, grâce aux marqueurs génétiquesgénétiques, de retracer les migrations préhistoriques de l'Homo sapiensHomo sapiens.


La méthode du projet génographique
Pour obtenir des résultats statistiques satisfaisants, le projet génographique prévoyait de recueillir, sur une durée de cinq ans, plus de 100.000 prélèvements d'ADN auprès de donneurs volontaires du monde entier. Les échantillons ont été analysés et comparés afin d'identifier les empreintes génétiquesempreintes génétiques de chaque individu. Elles ont été « regroupées » ensuite par type afin d'établir une carte mondiale des branches humaines... et pour dans un deuxième temps, tenter de retrouver les mouvements de population depuis 60.000 ans.
Le 13 avril 2005, National Geographic Society et IBMIBM ont lancé cette recherche révolutionnaire visant à retracer l'histoire des migrations de l'espèceespèce humaine. Le projet génographique, sous la direction de M. Spencer Wells, Ph.D. appuyé de façon généreuse par la Waitt Family Foundation, sera conservé dans un musée virtuel de l'histoire de l'humanité. L'intérêt de cette base de donnéesbase de données ? Héberger l'une des plus importantes collections d'information génétique jamais regroupées auparavant et constituer une ressource sans précédent pour les généticiensgénéticiens, les historienshistoriens et les anthropologues.
Les trois composantes clés du projet :
- Étude sur le terrain : La collecte d'échantillons d'ADN et la recherche sur le terrain auprès des populations autochtones sont les composantes clés du projet. Un conseil consultatif international supervise la sélection des populations autochtones à tester et assure le respect des protocolesprotocoles de l'échantillonnageéchantillonnage et de l'étude.
- Campagne de participation et de sensibilisation du public : Les particuliers peuvent participer au projet ce qui leur permettra de suivre la progression générale du projet tout en découvrant l'histoire de leur propre migration. Ces résultats personnels seront consignés de façon sécuritaire et anonyme pour assurer le respect de la vie privée des participants.
- Projet commémoratif : Les recettes de la vente des trousses de participation au projet contribueront au financement de futures études d'un projet fondé sur l'intérêt porté aux cultures du monde. Ce projet appuiera des projets d'éducation et de préservation auprès des groupes autochtones participants.
Le projet génographique s'est terminé en mai 2019.
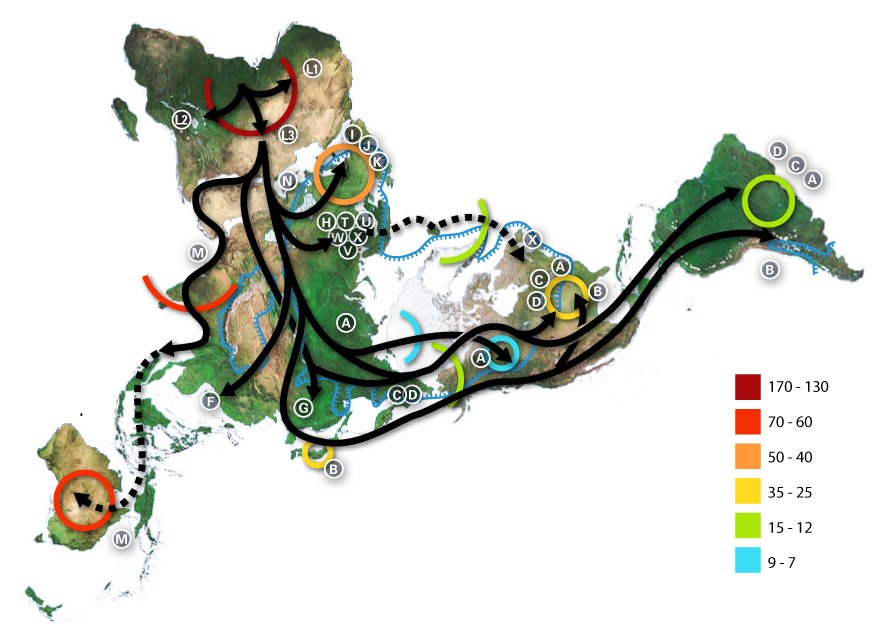
. <em>Africa h</em><em>arboring the start of the migration, is at the top left and South America at the far right. Migration patterns are based on studies of mitochondrial (matrilinear) DNA. Dashed lines are hypothetical migrations. Numbers represent thousand years <a title=\"w:before present\" href=\"https://en.wikipedia.org/wiki/before_present\" target=\"_blank\">before present</a>. The blue line represents area covered in <a title=\"w:ice\" href=\"https://en.wikipedia.org/wiki/ice\" target=\"_blank\">ice</a> or <a title=\"w:tundra\" href=\"https://en.wikipedia.org/wiki/tundra\" target=\"_blank\">tundra</a> during the last great ice age. The letters are the <a title=\"w:mitochondrial DNA\" href=\"https://en.wikipedia.org/wiki/mitochondrial_DNA\" target=\"_blank\">mitochondrial DNA</a> <a title=\"wikipedia:Haplogroup\" href=\"https://en.wikipedia.org/wiki/Haplogroup\" target=\"_blank\">haplogroups</a> (pure motherly lineages); Haplogroups can be used to define <a title=\"wikipedia:Genetic\" href=\"https://en.wikipedia.org/wiki/Genetic\" target=\"_blank\">genetic</a> populations and are often geographically oriented. For example, the following are common divisions for <a title=\"en:MtDNA haplogroup\" href=\"https://en.wikipedia.org/wiki/MtDNA_haplogroup\" target=\"_blank\">mtDNA haplogroups</a>: </em><em>African: L, L1, L2, L3 - </em><em>Near Eastern: J, N. </em><em>The dashed path for X, stopping in Iceland, gestures at the \"Solutrean hypothesis\", an idea discussed in the popular media at the time the map was made but mostly discredited since. </em><em>Southern European: J, K - </em><em>General European: H, V - </em><em>Northern European: T, U, X - </em><em>Asian: A, B, C, D, E, F, G (note: M is composed of C, D, E, and G) - </em><em>Native American: A, B, C, D, and sometimes X - The letters I, J, K shown in Europe are apparently included by mistake, as they refer to Y-DNA haplogroups. </em>© Avsa, <em>Wikimedia commons</em>, CC 2.5")
La génétique, science de l'hérédité
La génétique des populations traite des fluctuations des fréquences des différents allèlesallèles d'un même gènegène au cours du temps dans une ou plusieurs populations, sous l'influence de la sélection naturellesélection naturelle, de la dérive génétiquedérive génétique, des mutations et des migrations, et cherche à expliquer l'adaptation et la spéciationspéciation. Cette application a donné naissance au néodarwinisme. Elle permet de comprendre les mécanismes de conservation et/ou de disparition des populations et/ou des espèces. La population est un ensemble d'individus qui montre une unité de reproduction : tous les individus ont la même probabilité de se croiser, mais se reproduisent moins ou pas avec les populations voisines. Une population est déterminée par des critères spatiaux, temporels et par un génomegénome collectif, somme de génotypesgénotypes individuels : ce n'est pas une espèce. Cette population est un modèle, elle ne correspond que très rarement à la réalité.


Notion de population théorique idéale et loi de Hardy-Weinberg
La prévision de la variabilité génétique d'une population est très difficile à réaliser du fait des mutations, de la transmission simultanée de plusieurs gènes etc. Pour éviter ces problèmes, on a fabriqué un modèle idéal et pratique pour les nombreuses études mathématiques de la génétique des populations : les fréquences des allèles et des génotypes suivent la loi de Hardy-Weinberg, modèle de référence en génétique des populations. Cette loi énonce que les fréquences alléliquesfréquences alléliques et les fréquences génotypiques restent stables de génération en génération. Ce n'est qu'un modèle !
Le polymorphisme : dans une population, on dit qu'un gène est polymorphe, s'il possède au moins deux allèles ayant une fréquence supérieure ou égale à 1 %. Sinon, si le gène existe quand même en plusieurs exemplaires, il est polyallélique. Un gène polymorphe est obligatoirement polyallélique. La variabilité génétique est le résultat des mutations qui font apparaître de nouveaux allèles.
Les mutations, l'effet fondateur, la dérive génétique et les pressions de sélection variables conduisent à des différences génétiques de plus en plus importantes.
Si les migrations sont l'occasion de transmission d'allèles d'une population à l'autre, la dérive génétique et la sélection provoquent des variations de fréquence des allèles de façon interne aux populations. Ce modèle se justifie intuitivement : il est clair que tous les individus d'une génération ne vont pas produire le même nombre de descendants; certains n'auront pas d'enfants (ne transmettent aucun gène), d'autres vont avoir une famille nombreuse et transmettre plusieurs copies de leurs gènes à la génération suivante.
Si la population est grande, la perte d'une copie d'un certain allèle présent chez un individu sans descendance sera compensée par le fait qu'un autre individu possédant le même allèle aura plusieurs descendants. De cette manière, les fréquences alléliques fluctuent peu dans les grandes populations.

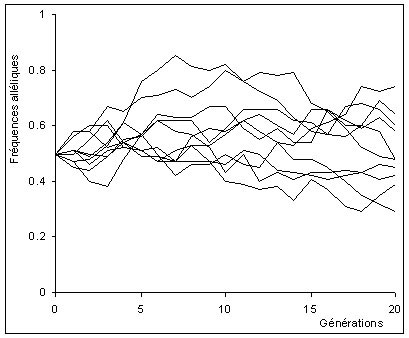
La dérive est de plus faible amplitude dans cette population. Les fréquences alléliques des populations restent relativement semblables entre elles et proches de la fréquence initiale de 0.5.
Par contre, si la population est petite, elles varient fortement car cet effet de moyenne n'agit pas. Plus rigoureusement, la variance de la fréquence allélique est inversement proportionnelle à la taille de la population.

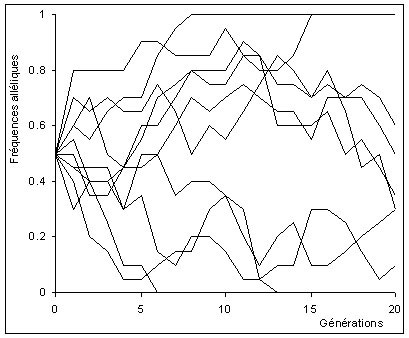
La dérive est de forte amplitude dans ces très petites populations. Elles divergent rapidement les unes des autres. Des allèles sont perdus et d'autres restent polymorphes ou se fixent. Donc, dans une grande population il y aura moins de fluctuations de fréquences d'une génération à l'autre. Il faut enfin faire attention au fait que la dynamique de la dérive génétique dépend de la taille efficace de la population et non de sa taille de recensement.
Conséquences de la dérive génétique
- La consanguinitéconsanguinité de la population va augmenter.
- Le nombre d'individus homozygoteshomozygotes va aussi augmenter.
- Les fréquences alléliques vont fluctuer de génération en génération
- L'amplitude de ces fluctuations sera d'autant plus grande que la population sera petite.
- Des populations issues d'une population mère vont se différencier génétiquement
Considérons que deux populations se séparent à un moment donné. Elles auront initialement les mêmes fréquences alléliques, mais elles vont connaître des processus de dérive génétique indépendants. Chacune des populations va avoir des fréquences alléliques se modifiant aléatoirement au cours du temps. Elles vont donc diverger génétiquement. La variabilité génétique de l'espèce va augmenter, ceci d'autant plus qu'elle sera composée de petites populations isolées les unes des autres. La fréquence allélique d'un groupe migrant n'est souvent pas représentative de la population dont il est issu. C'est l'effet fondateur.

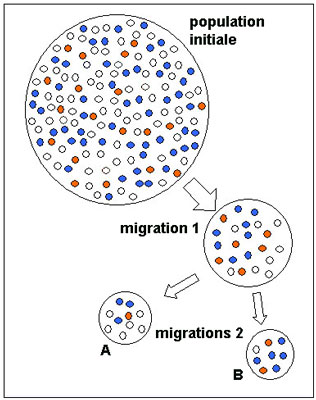
Il y a trois gènes présents dans la population de départ de notre exemple avec beaucoup de blancs, un peu moins de bleus et peu de rouges. Après une première migration les gènes blancs et bleus sont représentés à égalité mais les gènes rouges sont surreprésentés, les migrants ayant « beaucoup » ce gène, ceux qui sont restés moins. On voit lors des deux migrations suivantes que la répartition change encore... on voit bien que les humains qui sont partis ont pris certains gènes, en ont laissé d'autres....et certains gènes, rares, peuvent avoir disparu. Ainsi le groupe sanguin B très présent en Asie, terre d'origine des petites populations de chasseurs-cueilleurschasseurs-cueilleurs qui ont franchi la Béringie pour coloniser l'Amérique, est presque inexistant dans les populations Amérindiennes actuelles : perdu en route sans doute !
Malgré tous ces aléas n'importe quel homme peut avoir des descendants avec n'importe quelle femme de la planète et leurs descendants seront féconds, on le voit tous les jours, donc nous sommes toujours de la même seule et unique espèce Homo sapiens sapiens, c'est la définition même de l'espèce !
Nos ancêtres communs
Comme on l'a vu au chapitre précédent il semble bien admis, par la communauté scientifique en tous les cas, que nous ayons tous des ancêtres communsancêtres communs en Afrique. Chacun de nous a 2 parents qui avaient eux-mêmes chacun 2 parents etc. soit 2n... ancêtres. Considérons le temps maintenant : vers l'an 1200 soit il y a 33 générations, chacun de nous avait 233 ancêtres soit plus de 8 milliards... Or la population mondiale atteignait avec peine son premier milliard vers 1830... Il faut donc bien diminuer ce nombre d'aïeux donc augmenter notre parenté d'autant et on voit bien que nous devons, mathématiquement, être tous parents ! D'autant que, plus on recule dans le temps, moins il y avait de monde donc plus on a d'ancêtres communs... Donc, tous nos gènes sont des copies des gènes de nos ancêtres communs, des premiers Hommes. Et mille siècles est un délai trop court pour que de nombreuses et importantes modifications soient survenues.
- La couleurcouleur de la peau : une simple question de gènes régulateurs.
Nous possédons tous des mélanocytesmélanocytes, produisant de la mélanine (pigmentpigment naturel) sous contrôle génétique des mêmes gènes. Les mélanocytes sont des cellules de la peau qui comportent des prolongements et qui contiennent des mélanosomesmélanosomes, organelles cellulaires qui, elles, contiennent de la mélaninemélanine qui se présente sous deux formes chez l'Homme, les deux formes étant présentes dans tous les cas mais dans des proportions différentes :
- les granules d'eumélanine, ronds et tendres se trouvent en plus grand nombre dans les peaux noires et jaunes ;
- les granules de phaeomélanine, plus irréguliers, présents en plus grand nombre dans les peaux claires surtout chez les rouquins.
C'est la quantité de mélanine produite dans les mélanocytes qui fonce plus ou moins notre épiderme. Parallèlement, la quantité et l'intensité des rayons solaires influent sur notre corps qui produit plus ou moins de mélanine : c'est le bronzage.

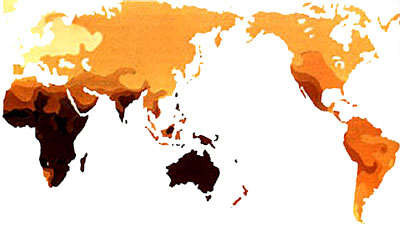
Les populations exposées de façon continue au soleilsoleil développent un bronzage permanent. D'une région tropicale vers le nord, vous rencontrez des populations de plus en plus claires, sans rupture. Il est impossible de déterminer à partir de quel moment un individu est blanc, noir ou jaune car toutes les nuances sont présentes.
C'est vrai qu'en Europe, par exemple, un Sicilien est plus brun qu'un Suédois si on les place l'un à côté de l'autre, mais, entre les deux on a le Français de Marseille, puis le Lyonnais, puis le Parisien, puis le Belge, puis l'Anglais etc. Sans compter que c'est sûrement pareil, pour les Chinois, entre la Chine du Sud et la Chine du Nord...





















, a été retrouvé dans l'eau du robinet d'une large majorité des villes où il a été recherché. © JLCo Ana Suanes, Adobe Stock")


