








au sommaire
Par leur diversité de formes et de couleurs, les micro-organismesmicro-organismes qui pullulent dans le planctonplancton marin offrent toujours de nouveaux sujets d'intérêt (curiosité intellectuelle) et d'émerveillement (surprise esthétique). Parmi eux, les radiolairesradiolaires sont assurément les plus élégants.


Les fines coques treillissées des radiolaires combinent presque à l'infini la sphère, le cylindre, le cônecône, le disque, l'anneau, la spirale... Et pourtant, si élégants soient-ils, les radiolaires comptent parmi les protistes marins les moins bien connus du biologiste. Cette méconnaissance résulte à la fois de leur faible dimension (de 50 à 300 µm en moyenne), de la complexité de leur organisation et de la difficulté de les maintenir en élevage. Pendant longtemps les paléontologuespaléontologues se sont peu intéressés à leurs tests car, même s'ils étaient connus dans des roches du PaléozoïquePaléozoïque, leur utilité stratigraphique était entachée d'une solide réputation d'inefficacité et leur extraction de la roche semblait délicate, voire impossible.
Caractères généraux des principaux groupes de radiolaires
Les radiolaires appartiennent au règne des protistes. Les protistes réunissent toutes les formes unicellulaires eucaryoteseucaryotes : protozoairesprotozoaires, alguesalgues et champignonschampignons. Les radiolaires sont des protozoaires, appartenant à la classe des actinopodes. Ils se caractérisent par la présence d'une capsule centrale séparant physiquement un ectoplasme d'un endoplasme. Leur classification est encore l'objet de débats.
Les acanthaires possèdent une capsule centrale percée de nombreux petits pores, des algues symbiotiques et un squelette en célestite (sulfate de strontium) très soluble dans l'eau. Les phéodaires présentent une capsule centrale percée de trois pores seulement, n'ont pas d'algues symbiotiques mais ont un pigment granulairegranulaire marron (= phaeodium). Les polycystines, dont la capsule centrale est perforée sur toute sa surface ou à une extrémité, possèdent des algues symbiotiques et un squelette de silicesilice intra- ou extra-capsulaire.
Pour certains auteurs, seuls les deux derniers groupes appartiennent aux radiolaires (Anderson, 1983). Les radiolaires polycystines sont les plus fréquemment préservés à l'état fossilefossile, aussi seul ce groupe est utilisé par les paléontologues.
Organisation cellulaire
Il existe une relation entre la capsule centrale (base de la classification des biologistes), qui différencie les radiolaires des autres actinopodes, et le squelette (base de la classification des paléontologues). Il existe donc des liens entre ces deux types de classification.
La grande complexité des radiolaires et la difficulté de comprendre leur organisation sont telles que le plus célèbre des précurseurs, Haeckel, a longtemps cru que ces organismes étaient multicellulaires. Hertwig (1876, 1879) a prouvé leur caractère unicellulaire et mis en évidence leur association symbiotique avec des algues.
Les radiolaires, tous marins, vivent pour la plupart à l'état d'individus isolés. Il existe cependant des formes coloniales, pouvant atteindre une grande taille (jusqu'à 5 mètres), comportant de très nombreux individus noyés au sein d'une masse gélatineuse mais où chacun possède généralement son propre squelette.
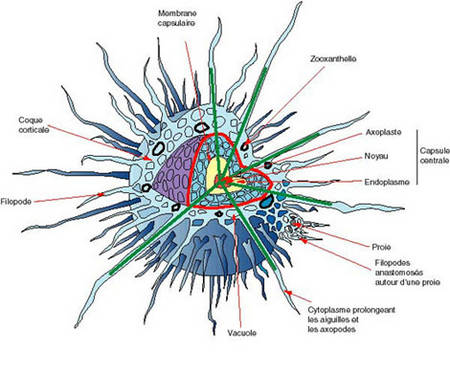
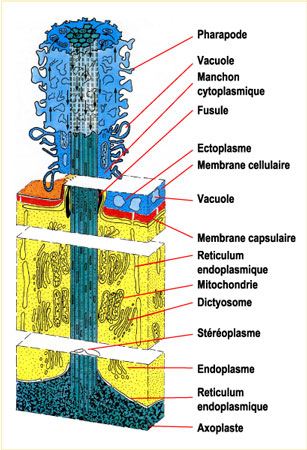
Les formes isolées ou les éléments unitaires des colonies offrent des formes très variées, mais présentent toujours un corps protoplasmique séparé en deux parties, ecto- et endoplasme, par une membrane capsulaire caractéristique de ce groupe. Le protoplasme est maintenu et orienté par un squelette (ou test).
La capsule centrale du plancton siliceux
La capsule centrale comprend l'endoplasme, le (ou les) noyau(x) et l'axoplaste ; elle est limitée par la membrane capsulaire.

Celle-ci est sphérique, ovale ou lobée. Sa forme diffère pour chaque espèceespèce. Elle croît avec l'âge, les plus grandes peuvent atteindre 400 µm.
. © N. Swanberg")
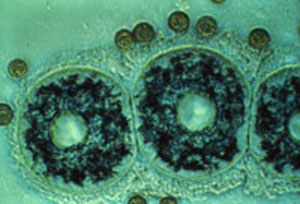
Le noyau est bien visible à l'intérieur d'une capsule centrale. La membrane capsulaire est ici clairement individualisée.
La membrane capsulaire : la capsule centrale est limitée par une membrane généralement pigmentée et bien visible sur les individus vivants. Sa couleur vive varie selon les espèces : rouge, jaune, marron, bleu-violet, vert... La membrane capsulaire est chitineuse ou pseudochitineuse, perforée par des fusules et fissures capsulaires.
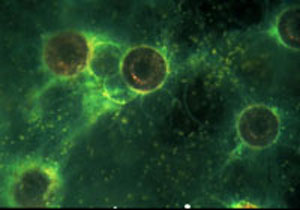
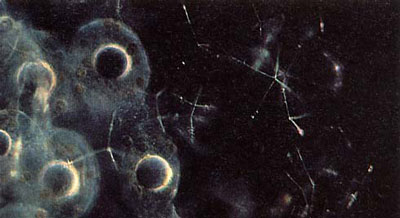
. On distingue bien la capsule centrale rougeâtre et le noyau des radiolaires. Les nombreuses zooxanthelles jaunâtres sont éparses dans l'ectoplasme de la colonie pour profiter au mieux de la lumière. © N. Swanberg")
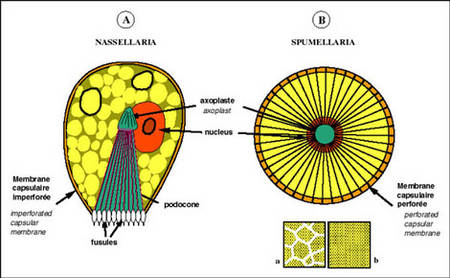
La membrane capsulaire est perforée et les pores sont disposés selon deux types principaux permettant ainsi de distinguer au sein des polycystines : les entactiniaires, les nassellaires et les spumellaires.

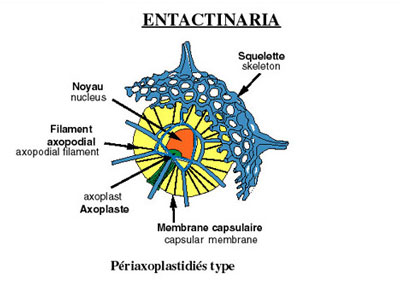
Chez les nassellaires et entactiniaires, les pores sont concentrés sur un seul secteur de la capsule. Les grandes fusules sont disposées en cône: le podocône.

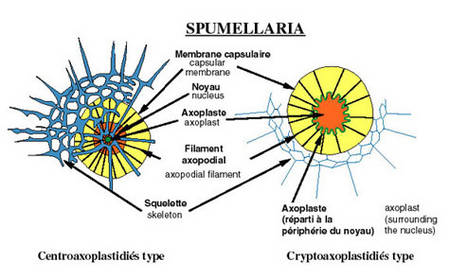
Chez les spumellaires, les pores de la membrane sont très nombreux et répartis sur toute la surface ou sous forme de pores regroupés dans des champs polygonaux. Le terme spumellaire est dû à la forme généralement sphérique du squelette.

L'endoplasme contient :
- un ou plusieurs noyaux, central(aux) ou excentré(s) ;
- les mitochondriesmitochondries, l'appareil de Golgiappareil de Golgi, des vacuolesvacuoles, des gouttelettes lipidiques, de composition variée, et des cristaux dont certains sont présumés être des protéinesprotéines. On sait aussi (Anderson & Matsuoka, 1992) que le cytoplasmecytoplasme intracapsulaire contient des bactériesbactéries et des algues ;
- l'axoplaste, organiteorganite d'où divergent les axopodes. Les axopodes sont longs, ténus et rigides, faits d'un filament axialaxial stéréoplasmique revêtu d'une gaine ectoplasmique, susceptibles de se rétracter, sous l'effet d'un choc par exemple.
 fait ressortir ses axopodes irisés à deux extrémités. La capsule centrale est bien visible ainsi que quatre épines. © N. Swanberg")
L'axoplaste joue un rôle lors de la formation du squelette ; en conséquence la morphologiemorphologie de celui-là guide celle du squelette.
Le corps extracapsulaire du plancton siliceux
Le corps extracapsulaire comprend : l'ectoplasme et ses inclusions et les pseudopodes. Ce vocable regroupe les filopodes, les axopodes et, parfois, l'axoflagelle.
L'ectoplasme : il forme un ensemble complexe d'aspect spongieux en coupe pouvant se rétracter, lors d'un choc par exemple, en une mince pellicule plaquée contre la membrane capsulaire. Lorsqu'il est épanoui, la couche formée par l'ectoplasme peut être subdivisée en trois zones qui sont, de l'intérieur vers l'extérieur.
 la sarcomatrice, fine, zone interne, de cytoplasme granuleux, pigments, en connexion avec la partie externe de l'endoplasme au travers des fusules. Les pseudopodes s'individualisent aux dépens de cette zone. (2) la calymma, zone médiane formée de cytoplasme gélatineux, caractérisée par l'abondance de vacuoles digestives et d'alvéoles (vacuoles ouvertes sur le milieu extérieur). Cette calymma est souvent colorée par les inclusions et symbiontes (zooxanthelles). (3) le sarcodictyum, couche fine de cytoplasme réticulé collé à la paroi externe.")
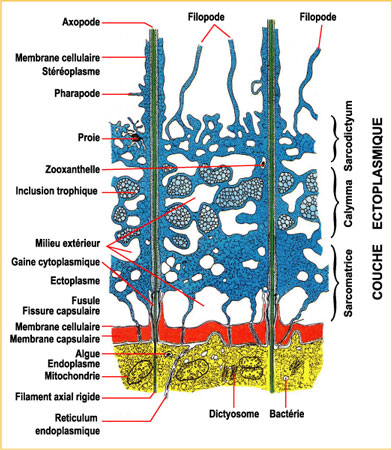
Les pseudopodes : ils correspondent à tous les tractus cytoplasmiques baignant dans le milieu extérieur. Ce vocable regroupe les filopodes, les axopodes et, parfois, l'axoflagelle. Les filopodes prennent naissance dans le sarcodictyum. Ce sont des extensions fines de cytoplasme, leur base peut être anastomosée. Ils servent à la capture des proies qu'ils entourent puis englobent lors de la digestiondigestion. Les axopodes prennent naissance sur l'axoplaste. Ils sont traversés par un filament axial rigide axoplasmique et enrobés par du cytoplasme mouvant.

Ce sont des organites tactiles et moteurs. L'axoflagelle est formé par la fusionfusion de plusieurs axopodes. C'est un organite tactile. Comme pour la couche ectoplasmique, les pseudopodes sont susceptibles de se rétracter contre la membrane capsulaire lors d'un choc ou d'un contact brutal, puis de s'épanouir de nouveau (Hollande & Enjumet, 1960).
Le squelette du plancton siliceux
C'est le seul élément d'étude pour le micropaléontologiste. Il est appelé indifféremment squelette, coque, test. En général, le test est inclus dans la couche ectoplasmique. Les épines elles-mêmes sont enrobées par des pseudopodes, auxquels elles servent d'axe. Ainsi, les parties siliceuses ne sont pas au contact direct de l'eau de mer et le squelette des organismes n'est pas sujet à la dissolution du vivant de la cellule. Le squelette des radiolaires s.l. est constitué le plus souvent de silice, empruntée au milieu marin et fixée par le protiste.

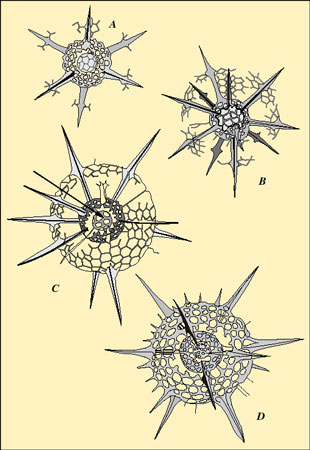
Le squelette des radiolaires est construit selon un plan géométrique bien défini, propre à chaque espèce. Il est donc important de connaître sa formation et son origine.Les coques ne se forment qu'à un moment bien précis de la croissance de l'organisme. Le dépôt de silice est un phénomène rapide. La croissance s'effectue de façon discontinue. Les périodes de fixation de silice résultent plus de la physiologie propre de l'organisme que de l'environnement. En outre, la croissance du squelette requiert parfois (sinon toujours) des fenêtresfenêtres précises de température mais supporte des variations importantes de salinitésalinité (Matsuoka & Anderson, 1992). Le squelette est fait d'une ou de plusieurs coques siliceuses (sphériques, concentriques, coniques...) d'où rayonnent éventuellement des épines. L'architecture du squelette peut être très simple (un seul spicule) ou très complexe. Les spumellaires (à gauche) et les nassellaires (à droite) se différencient, en général, par la morphologie de leur squelette.

Le squelette des spumellaires est généralement sphérique et porteporte des épines rayonnantes. Il comporte souvent plusieurs coques emboîtées, connectées par des barres radiaires d'épaisseur variable, se correspondant ou non d'une coque à l'autre. Celui des nassellaires est caractérisé, extérieurement, par une symétrie axialeaxiale (ex. formes coniques) et, intérieurement, par un spicule initial.
Rares sont les radiolaires actuels dépourvus de squelette. Encore est-il possible que ces formes soient des stades juvéniles. La coque est incluse dans le cytoplasme, les parties les plus internes pouvant être dans l'endoplasme ou même dans le noyau (Hollande & Enjumet, 1960). En général cependant, le test est inclus dans la couche ectoplasmique. Les épines elles-mêmes sont enrobées par des pseudopodes, auxquels elles servent d'axe. Ainsi, les parties siliceuses ne sont pas au contact direct de l'eau de mer, et le squelette des organismes n'est pas sujet à la dissolution du vivant de la cellule.
. </em>On distingue les capsules centrales et leur noyau interne entre lesquels sont dispersées des algues symbiotiques : les zooxanthelles (en vert-jaune) et des squelettes siliceux - partie droite de la photo, spicules tétraédriques. Diamètre des capsules centrales : 20 µm environ. © N. Swanberg")
Origine et mode de sécrétion du squelette
Le squelette des radiolaires est construit selon un plan géométrique bien défini, propre à chaque espèce. Il est donc important de connaître sa formation et son origine. Le test des radiolaires est relativement complexe. Les études ayant abordé sa formation ont été conduites à partir de divers points de vue, soit essentiellement physiquephysique (morphologie mathématique), soit biologique. Les apports sont alors radicalement différents.
Approche physique
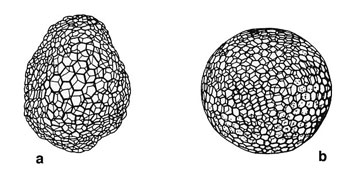
Selon Thompson (1942), la géométrie du squelette répond aux mêmes lois de physique fondamentale que celles qui régissent les interfaces entre fluides. Cet auteur a comparé la géométrie du squelette des radiolaires à différents modèles physiquesmodèles physiques applicables aux traits microstructuraux et macrostructuraux.
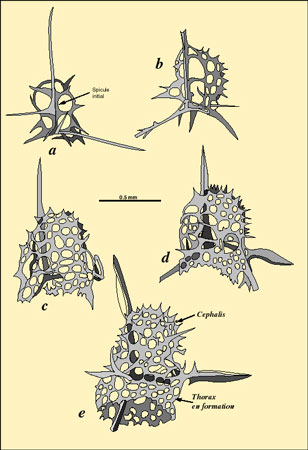
Il attira l'attention sur la similitude existant entre le modèle hexagonal de base de la coque treillissée de certains radiolaires et le réseau hexagonal visible sur certains tissus artificiels produits par adsorptionadsorption différentielle (Fig. a et b). Il montre aussi que le tétraèdre curviligne, obtenu par interaction de différentes bulles de savon dans un espace confiné est comparable à la forme de base de quelques squelettes de Nassellaires (Fig. c à e).
. a : Structure d'un réticulum plasmatique résultat de tensions superficielles. b : Structure d'un squelette de Radiolaire (<em>Auloniahexagona</em>).")

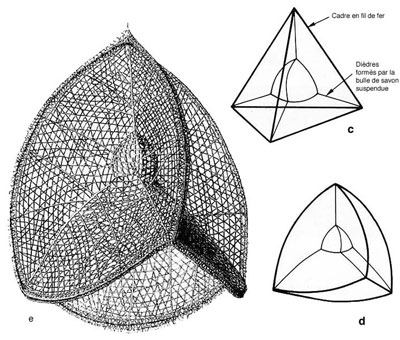
Thompson (1942) imagine que la pellicule cytoplasmique présente des vésicules juxtaposées d'une façon donnée. Celles-ci, par suite de tensions superficielles, deviennent prismatiques et en leur point de jonction la silice se déposerait selon certaines arêtes communes des prismes pour donner un réseau à mailles hexagonales. Si l'on a, à la place d'alvéoles polyédriques, un réseau de canaliculescanalicules anastomosées, on verra se constituer un réseau spongieux. Reprenant l'idée de Thompson, (op. cit.), Schaaf (1981) a montré, à l'aide de modèles mathématiques, qu'au cours du CénozoïqueCénozoïque, l'évolution de la morphologie des radiolaires tend vers une diminution des besoins en silice pour créer le test (régularisation de la forme, disposition des pores).
Cette idée d'économie en silice avait été proposée par Moore (1969) pour les radiolaires de l'Éocène au QuaternaireQuaternaire, les plus récents étant quatre fois plus légers, en moyenne, que les plus anciens. Selon Harper & Knoll (1975), la diminution pondérale du squelette des radiolaires, observée au cours du Cénozoïque, résulte d'une pressionpression sélective exercée par l'entrée des diatoméesdiatomées dans le cycle de la silice depuis la fin du MésozoïqueMésozoïque, le budget en silice dans l'eau étant supposé stable, voire diminuant dans l'océan. On notera encore que l'observation de certaines moléculesmolécules de fullerènesfullerènes - qui ne doivent rien à un quelconque cytoplasme - évoquent largement des radiolaires, de même que certaines fractales. Il y a donc clairement une influence de contraintes externes physiques sur la forme des radiolaires. Le squelette résulte donc de phénomènes purement physiques. Si ceux-ci sont les seules contraintes, il n'y a aucune raison pour que les différences morphologiques correspondent à des différenciations spécifiques et puissent induire des évolutions phylétiques. Les formes ne seraient que des produits du hasard.
Approche biologique
Anderson (1986) insiste sur le fait que la morphologie parfois très complexe de certains squelettes peut difficilement être décrite et expliquée par de simples réactions d'interfaces. Enriques (1931) avait supposé que le dépôt de silice était dépendant d'un moule organique. On sait aujourd'hui qu'il existe une matrice granuleuse organique précédant la silicification (Anderson, 1986).
. Un manchon granuleux se forme et précède une épine en silice reconstruite en une heure. Les traits irisés correspondent à des axopodes (caractéristiques des radiolaires). © N. Swanberg")
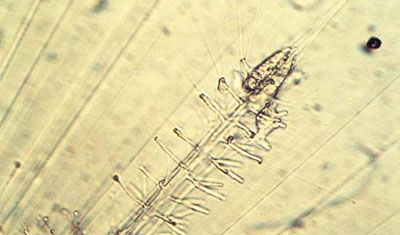
La distribution et la densité des granules à l'intérieur du squelette influencent d'ailleurs sa porositéporosité, sa résistancerésistance à la dissolution et donc ensuite ses transformations diagénétiques, c'est-à-dire sa conservation ou non dans les sédimentssédiments (Anderson & Swanberg, 1981). Le squelette est induit par une organisation du moule organique spécifique à l'espèce, il résulte d'une nécessité. Le test est donc susceptible d'évoluer avec l'espèce vivante et possède ainsi une valeur phylétique. La trame ontogénique est alors du second type, et la croissance interne du premier type (= hasard et nécessité), ce qui explique qu'après certaines remises à zéro des compteurs (grandes crises du monde vivant), on retrouve des convergences de formes. Ce sont des réponses voisines liées aux contraintes externes, contraintes toujours les mêmes et aux possibilités internes limitées.
Croissance
Une connaissance exacte de la croissance du squelette présente un grand intérêt pour le taxonomiste, mais les données restent fragmentaires en raison des difficultés d'élevage des radiolaires. Ainsi l'identification des formes juvéniles et l'analyse de leur transformation vers les stades adultes font-elles toujours partie du domaine des recherches à entreprendre. L'observation de formes voisines suggère la présence chez quelques espèces d'individus juvéniles, intermédiaires et adultes. Le faible nombre de formes juvéniles rencontrées est dû, en partie, au fait que les filets de collecte, ou les tamis, ont habituellement une maille de 63 µm à travers laquelle les individus jeunes peuvent passer.
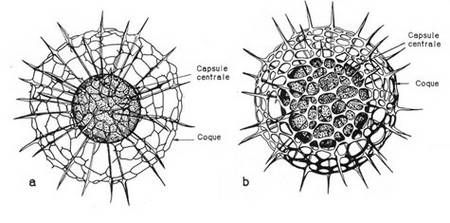
. Seul le squelette siliceux et la capsule centrale sont représenté. a : Forme jeune : le squelette n'est pas encore complètement formé, la capsule centrale est de relativement petite taille. b : Forme adulte : le squelette est terminé, le réseau de la coque s'épaissit et, corrélativement, la taille des pores diminue. La capsule centrale a augmenté de taille. Les coques ne se forment qu'à un moment bien précis de la croissance de l'organisme. Le dépôt de silice est un phénomène rapide. Il n'a en effet jamais été observé un stade pelliculaire ou partiellement silicifié.")
Deux processus de morphogenèsemorphogenèse sont connus : croissance par ponts et développement par dépôt de bordure (De Wever, 1981a ; Anderson, 1986). Les coques s'individualisent à partir d'épines. À l'origine, les éléments du squelette sont grêles, leur épaississement est dû à un dépôt additionnel de silice. Chez un même individu, une coque épaisse à pores circulaires, par exemple, succède à une coque grêle à mailles hexagonales et une épine tricarénée massive n'est tout d'abord qu'une baguette siliceuse ténue (Matsuoka, 1992a). Des épines, se développe un réseau de branches qui s'anastomosent. Or, les épines servent d'axe à des pseudopodes dont le nombre et la position sont fonction du nombre et de la position des fusules de la membrane capsulaire, caractéristiques de chaque espèce. La position de chacun d'eux est donc également fixe. Cette donnée essentielle souligne les rapports entre architecture cellulaire et organisation squelettique.
Chez les nassellaires, il existe des stades de croissance. On voit les spicules de base s'entourer progressivement d'épines ou parois. Chez les spumellaires, il fut supposé un moment que la croissance pouvait être centripète. On sait maintenant qu'elle est centrifuge. La coque la plus interne est la première à s'individualiser, probablement dans l'ectoplasme. La croissance s'effectue de façon discontinue puisque des phases de squelettogenèse rapides alternent avec des stases. Les périodes de fixation de silice résultent plus de la physiologie propre de l'organisme que de l'environnement (Matsuoka, 1992a).
En outre, la croissance du squelette requiert parfois (sinon toujours) des fenêtres précises de température mais supporte des variations importantes de salinité (Matsuaka & Anderson, 1992). Par suite de la croissance du volumevolume cytoplasmique et donc de sa capsule centrale, il est fréquent que celle-ci englobe la coque déjà formée. Le squelette dans certains cas baigne alors dans l'endoplasme, dans d'autres il reste entouré d'une fine pellicule d'ectoplasme, la capsule centrale étant lobée.
Structure du squelette
Le squelette est fait d'une ou de plusieurs coques siliceuses (sphériques, concentriques, coniques...) d'où rayonnent éventuellement des épines. Selon l'âge du protiste, les coques sont extérieures à la capsule centrale ou non, et sont alors incluses dans l'endoplasme ou même dans le noyau. On distingue généralement trois types de coques : la coque treillissée (ou grillagée), la coque spongieuse et la plaque perforée.
Une coque treillissée est constituée par un réseau de barres délimitant des pores très rapprochés. La forme de base des pores est généralement hexagonale mais le dépôt de silice à l'intérieur des pores peut les obstruer en partie et leur conférer un contour circulaire. On reconnaît des formes où le réseau polygonal primitif est visible et d'autres où il est estompé. La forme générale des pores et leur distribution sont constantes pour une espèce et sont donc utilisées pour la taxonomietaxonomie. Une coque spongieuse épaisse ou mince est un entrelacs irrégulier de trabécules longs ou courts, fins ou épais, dans un réseau tridimensionnel. Un modèle défini de pores ne peut alors pas être reconnu. Une coque en plaque perforée présente des petits pores très espacés sans arrangement géométrique discernable. Les pores des coques treillissées ou des plaques perforées sont circulaires, polygonaux ou irréguliers, égaux ou inégaux, régulièrement disposés ou non. Les épines sont circulaires en section transverse ou cannelées, triradiées ou tétraradiées. La présence d'épines tétraradiées est considérée comme étant un caractère primitifcaractère primitif (Dumitrica, 1982).



, il fait environ 5.900 °C. © Pascal Paquereau, Spaceweather")








